Veranderingen in de samenstelling van het menselijke microbioom ━ de verzameling van alle micro-organismen in en op ons lichaam ━ worden in verband gebracht met verschillende inflammatoire en (auto)immuunziekten. Hoewel de precieze rol van het microbioom bij het ontstaan en beloop van deze ziekten nog niet geheel duidelijk is, beginnen we steeds meer te begrijpen over de complexe interacties tussen de microben en hun gastheer en het belang hiervan voor de ontwikkeling en homeostase van het immuunsysteem.
Dit artikel beschrijft de huidige kennis over deze gastheer-microbe-interacties en de rol van verstoringen in het microbioom bij het ontstaan van ontstekingsziekten, in het bijzonder allergie en astma.
(NED TIJDSCHR LEEFSTIJLGENEESKD 2023;1:22–8)
Druk op de afspeelknop om de tekst voor te lezen.
SAMENVATTING
Veranderingen in de samenstelling van het menselijke microbioom ━ de verzameling van alle micro-organismen in en op ons lichaam ━ worden in verband gebracht met verschillende inflammatoire en (auto)immuunziekten. Hoewel de precieze rol van het microbioom bij het ontstaan en beloop van deze ziekten nog niet geheel duidelijk is, beginnen we steeds meer te begrijpen over de complexe interacties tussen de microben en hun gastheer en het belang hiervan voor de ontwikkeling en homeostase van het immuunsysteem.
Dit artikel beschrijft de huidige kennis over deze gastheer-microbe-interacties en de rol van verstoringen in het microbioom bij het ontstaan van ontstekingsziekten, in het bijzonder allergie en astma.
(NED TIJDSCHR LEEFSTIJLGENEESKD 2023;1:22–8)
INLEIDING
Het menselijk lichaam herbergt een complex ecosysteem van biljoenen bacteriën die samen met archaea (naast bacteriën een apart domein van eencellige prokaryoten), virussen, gisten en schimmels het ‘microbioom’ vormen. De huid, luchtwegen en urogenitaal stelsel herbergen alle hun eigen microbioom, maar veruit de meeste micro-organismen bevinden zich in het maagdarmkanaal. Dit darmmicrobioom speelt niet alleen een belangrijke rol in de spijsvertering en synthese van essentiële vitamines, maar ook in de bescherming tegen binnendringende ziekteverwekkers en de ontwikkeling en homeostase van het immuunsysteem. De eerste levensjaren zijn hierbij een cruciale periode waarin geleidelijk dit complexe ecosysteem van micro-organismen tot stand komt. Dit is tevens de belangrijkste periode waarin het microbioom de ontwikkeling van enterale mucosale weefsels en immuunsysteem stimuleert met zowel lokale als systemische effecten tot gevolg.1,2
In welvarende, geïndustrialiseerde landen is de mate en diversiteit van microbiële blootstelling sterk afgenomen. Hierdoor wijkt de ontwikkeling van het menselijke darmmicrobioom sterk af van het microbioom van onze voorouders.3,4 Dit heeft mogelijk mede bijgedragen aan de toename in allergieën en autoimmuunziekten, zoals de ‘oude vrienden’- of ‘biodiversiteitshypothese’ stelt.5 Deze verfijning van de klassieke ‘hygiënehypothese’ veronderstelt namelijk dat niet zozeer blootstelling aan acute infecties, maar blootstelling aan micro-organismen waarmee we zijn geëvolueerd (‘oude vrienden’) nodig is voor een adequate ontwikkeling van het immuunsysteem. Het betreft hier persisterende, subklinische infecties (zoals bepaalde parasitaire worminfecties), maar bovenal ook de kolonisatie van gunstige micro-organismen in het darmkanaal.6
ONTWIKKELING VAN HET DARMMICROBIOOM EN VERSTORINGEN DOOR LEEFSTIJLVERANDERINGEN
Hoewel sommige onderzoeken anders suggereren, is er vooralsnog onvoldoende wetenschappelijk bewijs voor het bestaan van een microbioom in een gezond foetaal milieu.7 Dit wil overigens niet zeggen dat prenatale microbiële blootstelling (o.a. via het maternale microbioom) geen rol speelt in de immunologische ontwikkeling van de foetus. Integendeel, dierstudies laten zien dat microbiële componenten en metabolieten overgedragen via de placenta een belangrijke rol kunnen spelen onder meer via epigenetische imprinting.8
Microbiële kolonisatie van zuigelingen lijkt echter pas aan te vangen zodra het amnionmembraan breekt. De manier van bevalling is hierbij bepalend voor de bacteriesoorten die als eerste arriveren, verlaagde aantallen bifidobacteriën en de nagenoeg volledige afwezigheid van Bacteroides bij kinderen geboren via een keizersnede zijn kenmerkend voor het gebrek aan transmissie van maternale vaginale, maar vooral fecale, bacteriën. De microbiële diversiteit neemt geleidelijk toe en facultatief anaerobe bacteriën, waaronder Enterobacteriaceae, streptokokken, stafylokokken, lactobacillen en enterokokken, worden gedurende de eerste weken tot maanden steeds verder teruggedrongen door obligaat anaeroben, zoals bifidobacteriën, Bacteroides, Veillonella en clostridia.9,10 Het type zuigelingenvoeding speelt hierbij een cruciale rol. Moedermelk bevat honderden verschillende, voor onze eigen spijsverteringsenzymen onverteerbare, complexe melksuikers (oligosacchariden). Bifidobacteriën zijn specialisten in het benutten van deze oligosacchariden en domineren dan ook de darmen van borstgevoede kinderen. Daarnaast bevat moedermelk vele andere bioactieve componenten en levende bacteriën die van invloed zijn op het zich ontwikkelende darmmicrobioom van zuigelingen. De introductie van vaste voeding en het stoppen van zuigelingenvoeding zorgen voor een afname van bifidobacteriën en Veillonella, een verdere toename van Bacteroides, terwijl Lachnospiraceae (o.a. Blautia en Roseburia) en Ruminococcaceae (o.a. Faecalibacterium, Ruminococcus) uitgroeien tot dominante bacteriesoorten.9,11–13 Veel van deze bacteriën produceren korte-ketenvetzuren (KKV’s), waaronder boterzuur, en de concentratie van deze metabolieten neemt dan ook sterk toe.14,15 Tot slot vestigen sommige prokaryoten, waaronder de archea Methanobrevibacter en bacteriesoorten Desulfovibrio, Bilophila en Christensenellaceae pas na de eerste levensjaren.16 Dit impliceert dat de juiste niche voor deze soorten pas later wordt gevormd. Veranderingen in leefstijl in de geïndustrialiseerde wereld hebben een overduidelijk effect gehad op het menselijke darmmicrobioom, met een verlies aan microbiële diversiteit en verdwijnen van specifieke bacteriesoorten tot gevolg.17,18 Zoals eerder aangegeven is een keizersnedebevalling een klassiek voorbeeld van de belemmering van overdracht van gunstige bacteriesoorten, maar ook kleinere gezinsgroottes, verminderd contact met de natuurlijke omgeving, het verlies van biodiversiteit in diezelfde omgeving, alsmede de verminderde inname van voedselgebonden bacteriën als gevolg van de toenemende consumptie van sterk bewerkt (fabrieks)voedsel, beperken alle de transmissiekans van mogelijke ‘oude vrienden’.19
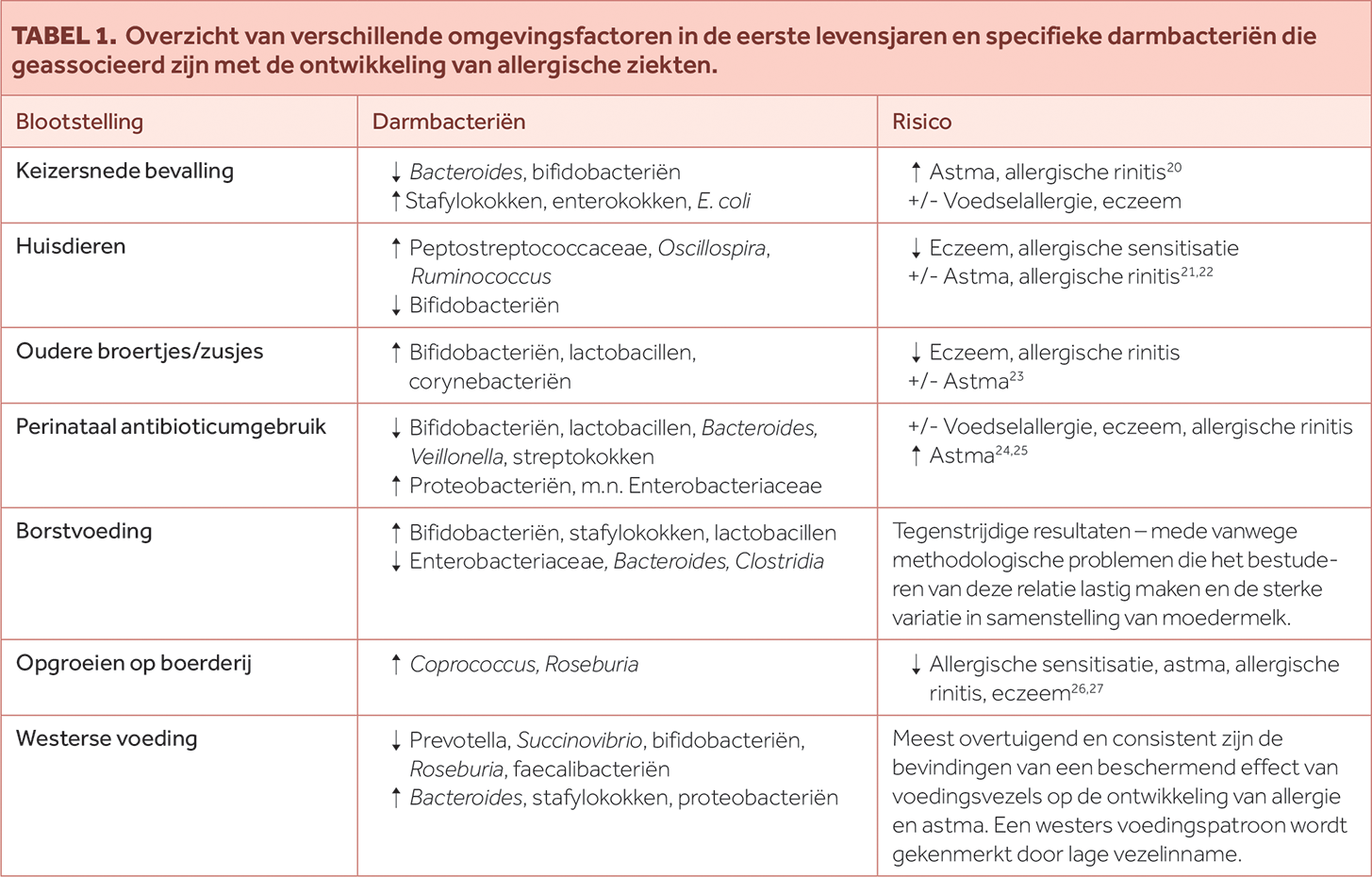
Daarnaast zijn er factoren die een selectieve druk uitoefenen op het darmmicrobioom. Hiertoe behoort het gebruik van antibiotica en andere medicatie, maar ook het westerse voedingspatroon (rijk aan dierlijke vetten, simpele suikers en arm in vezels) dat zorgt voor een verlaging in microbiële diversiteit en KKV-producerende bacteriesoorten (zie Tabel 1).
TABEL 1. Overzicht van verschillende omgevingsfactoren in de eerste levensjaren en specifieke darmbacteriën die geassocieerd zijn met de ontwikkeling van allergische ziekten.
POSTNATALE ONTWIKKELING VAN HET MUCOSALE IMMUUNSYSTEEM
Het mucosale imuunsysteem staat voor de belangrijke uitdaging om actief te reageren tegen pathogene micro-organismen, maar tegelijkertijd de commensale micro-organismen en voedselantigenen te tolereren om zo ongewenste immuunreacties tegen deze onschadelijke antigenen te voorkomen. Studies met kiemvrije dieren tonen aan hoe belangrijk het microbioom is voor de ontwikkeling en training van dit mucosale immuunsysteem. Kiemvrije dieren hebben een onderontwikkeld immuunsysteem met lagere productie van antimicrobiële eiwitten, minder intra-epitheliale lymfocyten, minder actieve Peyerse platen en lagere aantallen IgA-producerende plasmacellen.28 Introductie van microben in deze kiemvrije dieren kan, zeker wanneer dit in de neonatale periode gebeurt, een normale ontwikkeling van het gastro-intestinale immuunsysteem herstellen.
Een goed ontwikkelde darmbarrière is een belangrijke eerste verdedigingslijn en voorkomt overmatige microbiële stimulering van epitheel- en immuuncellen. Deze darmbarrière wordt mede gevormd door de occludensverbindingen (‘tight junctions’) tussen aangrenzende darmepitheelcellen, productie van een mucuslaag door mucine-producerende Goblet-cellen, secretie van antimicrobiële eiwitten (defensines, cathelicidines) door Panethen epitheelcellen en de productie van secretoir IgA. Microbiële kolonisatie speelt een belangrijke rol bij de ontwikkeling en regulatie van deze darmbarrière. Zo kan IL-18-secretie door epitheelcellen zowel direct door micro-organismen worden gestimuleerd, via stimulatie van patroonherkenningsreceptoren, als indirect door microbiële metabolieten als boterzuur. IL-18 kan vervolgens de barrièrefunctie bevorderen door de productie van antimicrobiële eiwitten en mucus te stimuleren.29
Ook indol, een afbraakproduct van tryptofaan, stimuleert de darmbarrièrefunctie. Deze bacteriële metaboliet activeert de aryl-hydrocarbonreceptor met verhoogde IL-22 door type 3 aangeboren lymfoïde cellen (ILC3), upregluatie van occludens en claudins en verhoogde mucusproductie tot gevolg.28–30
Verder zijn er aanwijzingen dat bepaalde darmbacteriën in staat zijn om, net als het menselijk lichaam, vitamine A en D om te zetten in de actieve metabolieten retinoïnezuur en calcitriol (1α,25-dihydroxyvitamine D3).31,32
Door binding aan de vitamine D- en retinoïnezuurreceptoren kunnen deze metabolieten de transcriptie van een breed scala aan genen beïnvloeden. Vitamine A en D bevorderen de barrièrefunctie door de productie van antimicrobiële eiwitten en expressie van ‘tight junctions’ te stimuleren.31,32 De interactie tussen vitamine A en D en het microbioom is complex en bidirectioneel, aangezien deze vitamines op hun beurt ook de samenstelling en diversiteit van het microbioom weer kunnen beïnvloeden.31
Dendritische cellen en regulatoire T-cellen (Tregs) spelen een belangrijke rol bij de ontwikkeling van immunologische tolerantie tegen onschadelijke antigenen. De ontwikkeling van immunologische tolerantie op jonge leeftijd loopt parallel met de ontwikkeling van de mucosale barrières en microbiële kolonisatie van diezelfde mucosale weefsels. Verstoringen in de ontwikkeling van het microbioom kunnen dan ook direct van invloed zijn op de (inadequate) programmering van immunologische tolerantie en het ontstaan van allergieën en auto-immuunziekten. Zo hebben vitamine A en D niet enkel een positief effect op de barrièrefunctie, maar ook op de ontwikkeling van immunologische tolerantie. De rol van retinoïnezuur in de ontwikkeling van Foxp3+ Tregs is welomschreven. Daarnaast hebben verschillende studies laten zien dat ook calcitriol een direct effect kan hebben op Tregs, maar ook de maturatie van dendritische cellen kan remmen en daarmee hun tolerogene eigenschappen behouden.33
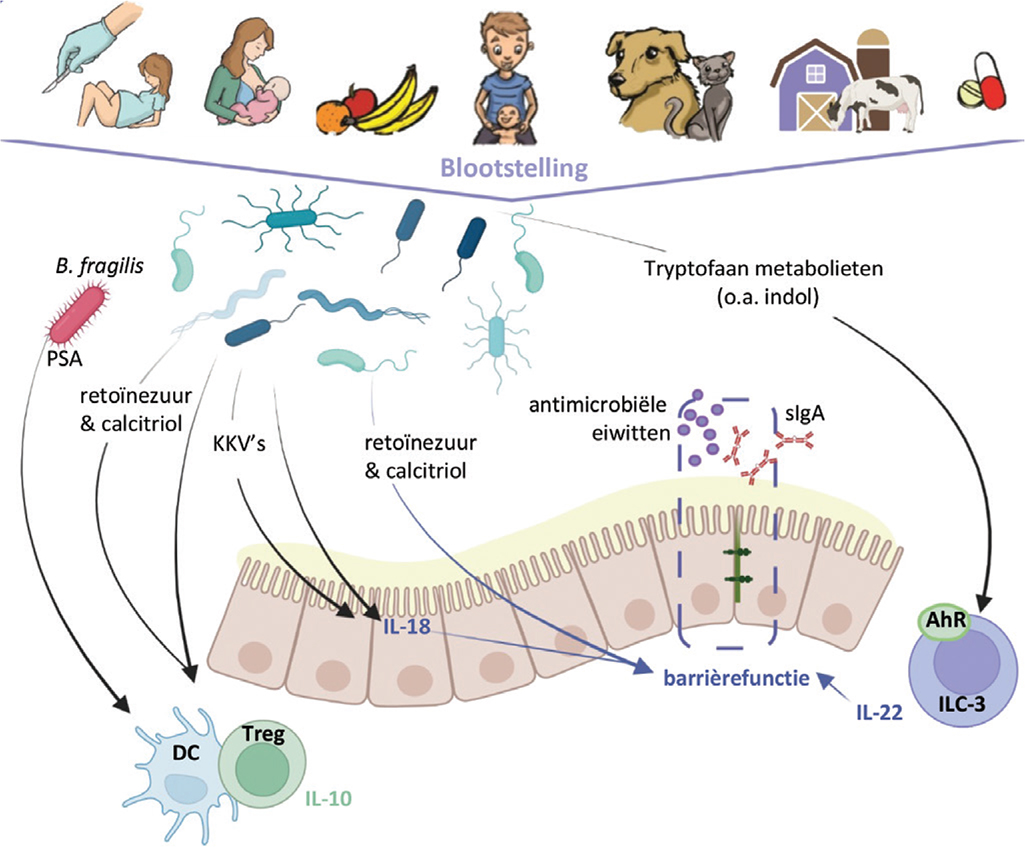
Polysaccharide A (PSA) geproduceerd door de darmbacterie Bacteroides fragilis is een goed voorbeeld van een microbiële structuur die Tregs kan induceren. Verschillende studies hebben aangetoond dat PSA, via TLR-2-afhankelijke activatie van plasmacytoïde dendritische cellen, IL-10-producerende Tregs kan induceren en daardoor kan beschermen tegen verschillende ontstekingsziekten.34 Ook meerdere soorten Clostridium blijken in staat de ontwikkeling van Foxp3+ Tregs te induceren, mogelijk via de productie van KKV’s.35 Dit zijn slechts enkele voorbeelden van specifieke bacteriën of bacteriële producten die van invloed zijn op het mucosale immuunsysteem, waarbij de resultaten veelal afkomstig zijn van in-vitro- of dierexperimenteel onderzoek. Het wordt steeds duidelijker dat het microbioom als een complex interactief netwerk opereert. Hoe dit geheel aan complexe interacties tussen het microbioom en de gastheer precies van invloed is op de immunologische ontwikkeling en regulatie in de mens is nog grotendeels onbekend (zie Figuur 1).
FIGUUR 1. Overzicht van belangrijke determinanten van het microbioom in de eerste levensjaren en van een aantal manieren waarop het microbioom en microbiële metabolieten het mucosale immuunsysteem beïnvloeden. Bron: Gecreëerd met Biorender.com AhR=aryl-hydrocarbonreceptor, B. fragilis=Bacteroides fragilis, DC=dendritische cellen, ILC-3=type 3 aangeboren lymfoïde cellen, KKV’s=korte-ketenvetzuren, PSA=polysaccharide A, sIgA=secretoir IgA.
DE ROL VAN HET MICROBIOOM IN DE ONTWIKKELING VAN ALLERGIE EN ASTMA
Inadequate tolerantie-inductie leidt in het geval van type 1 al lergische reacties tot een Th2-type immuunrespons, waarbij B-cellen worden aangezet tot de productie van IgE. Deze IgEantilichamen binden vervolgens aan mestcellen en bij een volgend contact met het allergeen zal ‘crosslinking’ van mestcelgebonden IgE plaatsvinden waarop mestcellen degranuleren. Dit leidt uiteindelijk tot de klinische symptomen bij de patiënt. Epidemiologische studies hebben de afgelopen jaren meermaals een associatie tussen het darmmicrobioom en allergische aandoeningen beschreven, waarbij longitudinale studies verder aantoonden dat verschillen in het microbioom voorafgaan aan de eerste allergische symptomen. Ondanks de heterogeniteit tussen studies in opzet, gebruikte microbiële technieken, klinische uitkomsten en de uiteindelijke resultaten, krijgen we langzaam inzicht in een aantal consistente patronen (zie Tabel 2).
TABEL 2. Overzicht van darmbacteriën die in meerdere studies geassocieerd bleken met de ontwikkeling van allergische ziekten.
Een groot aantal studies vond namelijk een verlaagde microbiële diversiteit gedurende het eerste levensjaar in relatie tot eczeem en allergische sensitisatie, terwijl een dergelijk verband voor astma vaak afwezig bleek.9,36–43 Daarnaast zijn er specifieke bacteriesoorten die verlaagd blijken bij kinderen die later allergie ontwikkelen in vergelijking tot kinderen die vrij blijven van allergische klachten. Het betreft dan vooral KKV-producerende bacteriën, waaronder Faecalibacterium (prausnitzii) en Anaerostipes, beide boterzuur-producerende bacteriën die geassocieerd zijn met een lager risico op voedselallergie, eczeem, piepen en astma.9,44–47 Ook acetaat-producerende bacteriën, zoals Lachnospira, lijken het risico op allergische aandoeningen te verlagen.9,46
In lijn met deze bevindingen liet de PASTURE-studie zien dat hogere concentraties van boter- en propionzuur in de ontlasting van 1-jarige kinderen geassocieerd was met een lager risico op allergische sensitisatie, voedselallergie, hooikoorts en astma op latere leeftijd.48
Bifidobacteriën zijn eveneens herhaaldelijk in verband gebracht met bescherming tegen allergische sensitisatie en klachten.47,49 Recentelijk werd aangetoond dat sommige soorten bifidobacteriën in staat zijn aromatische aminozuren, waaronder tryptofaan, om te zetten in aromatische melkzuren. Deze aromatische melkzuren stimuleren de aryl-hydrocarbonreceptor, een belangrijke receptor in de regulatie van darmhomeostase en immuunreacties.49
Het meest overtuigende bewijs voor een causale rol van het darmmicrobioom in de etiologie van allergische aandoeningen komt van dierexperimenteel onderzoek. Kiemvrije dieren vertonen overmatige IgE-productie en een ernstigere anafylaxe na orale sensitisatie dan conventionele dieren – een fenomeen dat omkeerbaar is door toediening van een beperkt aantal bacteriesoorten.50 Ook het verstoren van het microbioom door antibiotica in neonatale, maar niet in volwassen, muizen resulteerde in hogere IgE-niveaus, verhoogde vatbaarheid voor allergische luchtwegklachten en verlaagde regulatoire T-cellen in de darmen.51
Deze resultaten onderschrijven niet alleen het belang van microbiële stimulatie in het vroege leven, maar ook de communicatie tussen de darmen en longen (de darm-long-as). Deze darm-long-communicatie kan deels ook het beschermende effect van opgroeien op een boerderij op de ontwikkeling van astma verklaren. Depner et al. toonden aan dat blootstelling aan de boerderij-omgeving leidde tot een meer volwassen microbioom bij 1-jarige kinderen. Deze snellere maturatie van het microbioom, gekenmerkt door hogere aantallen van de boterzuurproducerende Roseburia en Coprococcus, kon een groot deel van het beschermende effect op astma verklaren.52
Microbiële metabolieten, zoals de KKV’s, spelen dus waarschijnlijk een belangrijke rol in deze communicatie, hetgeen verder wordt bevestigd door dierexperimentele studies. Zo toonden Thorburn et al. in een muismodel aan dat orale toediening van azijnzuur tot vermindering van de allergische ontstekingsreactie in de luchtwegen leidt.53 Hierbij bleek azijnzuur het aantal en de functie van regulatoire T-cellen te bevorderen via epigenetische veranderingen in de Foxp3-promotor.
CONCLUSIE
De afgelopen jaren heeft het onderzoek naar de complexe interacties tussen het darmmicrobioom en het immuunsysteem tot vele nieuwe inzichten geleid. Ondanks dat er ook nog vele vragen en uitdagingen zijn, is het belang van het microbioom in de ontwikkeling van het mucosale immuunsysteem inmiddels overduidelijk. In welke mate de veranderingen in het darmmicrobioom als gevolg van verminderde microbiële blootstelling (aan ‘oude vrienden’) heeft bijgedragen aan de toename in allergieën en auto-immuunziekten staat nog minder overtuigend vast. Dierexperimenteel onderzoek ondersteunt deze relatie en heeft verschillende mechanismen ontrafeld die de invloed van specifieke bacteriën en bacteriële metabolieten op de ontwikkeling van immunologische tolerantie verklaren. Humaan onderzoek is vooralsnog voornamelijk observationeel en beperkt zich meestal tot het karakteriseren van de samenstelling van de microbiota. Het integreren van andere ‘-omics’-technieken (‘metabolomics’, ‘metaproteomics’, ‘single-cell transcriptomics’) in toekomstig onderzoek kan bijdragen om de complexe interacties tussen het microbioom en het immuunsysteem verder te ontrafelen. Bovendien kunnen de inzichten die tot dusver zijn verkregen over de rol van specifieke bacteriën en bacteriële metabolieten bijdragen aan het ontwikkelen van interventiestrategieën gebaseerd op basis van bewijs, bijvoorbeeld in de vorm van toediening van specifieke bacteriële lysaten, consortia van levende micro-organismen, metabolieten als KKV’s of contact met (een divers microbioom) in de natuurlijke leefomgeving. Enkel op deze manier kunnen we een oorzakelijk verband vaststellen en de mogelijke gevolgen van het gemis aan onze ‘oude vrienden’ herstellen.
AANWIJZINGEN VOOR DE PRAKTIJK
Microbiële kolonisatie in het vroege leven speelt een belangrijke rol bij de ontwikkeling van zowel het aangeboren als verworven immuunsysteem.
De rol van korte-ketenvetzuren in de ontwikkeling van immunologische tolerantie onderstreept het belang van een vezelrijk voedingspatroon.
Toediening van specifieke bacteriële lysaten, consortia van levende micro-organismen en/of microbiële metabolieten biedt kansen om het immuunsysteem te versterken en daarmee de ontwikkeling van immuungemedieerde aandoeningen te voorkomen.
Referenties
1. Al Nabhani Z, Eberl G. Imprinting of the immune system by the microbiota early in life. Mucosal Immunol 2020;13:183–9. 2. Renz H, Adkins BD, Bartfeld S, et al. The neonatal window of opportunity-early priming for life. J Allergy Clin Immunol 2018;141:1212–4. 3. Blaser MJ, Falkow S. What are the consequences of the disappearing human microbiota? Nat Rev Microbiol 2009;7:887–94. 4. Sonnenburg ED, Sonnenburg JL. The ancestral and industrialized gut microbiota and implications for human health. Nat Rev Microbiol 2019;17:383–90. 5. Rook GA, Raison CL, Lowry CA. Microbial ‘old friends’, immunoregulation and socioeconomic status. Clin Exp Immunol 2014;177:1–12. 6. Savelkoul HF. [The hygiene hypothesis: a fruitful concept in immunological research]. Ned Tijdschr Geneeskd 2006;150:2596. 7. Hornef M, Penders J. Does a prenatal bacterial microbiota exist? Mucosal Immunol 2017;10:598–601. 8. Gomez de Aguero M, Ganal-Vonarburg SC, Fuhrer T, et al. The maternal microbiota drives early postnatal innate immune development. Science 2016;351:1296–302. 9. Galazzo G, van Best N, Bervoets L, et al. Development of the microbiota and associations with birth mode, diet, and atopic disorders in a longitudinal analysis of stool samples, collected from infancy through early childhood. Gastroenterology 2020;158:1584–96. 10. Penders J, Van Best N. The development of the gut microbiota in childhood and its distortion by lifestyle changes. In: Rook GA, Lowry CA, eds. Evolution, biodiversity and a reassessment of the hygiene hypothesis. Springer Cham 2022;89:197–219. 11. Nagpal R, Tsuji H, Takahashi T, et al. Ontogenesis of the gut microbiota composition in healthy, full-term, vaginally born and breast-fed infants over the first 3 years of life: a quantitative bird’s-eye view. Front Microbiol 2017;8:1388. 12. Podlesny D, Fricke WF. Strain inheritance and neonatal gut microbiota development: a meta-analysis. Int J Med Microbiol 2021;311:151483. 13. Yassour M, Vatanen T, Siljander H, et al. Natural history of the infant gut microbiome and impact of antibiotic treatment on bacterial strain diversity and stability. Sci Transl Med 2016;8:343ra81. 14. Differding MK, Benjamin-Neelon SE, Hoyo C, et al. Timing of complementary feeding is associated with gut microbiota diversity and composition and short chain fatty acid concentrations over the first year of life. BMC Microbiol 2020;20:56. 15. Nilsen M, Madelen Saunders C, Leena Angell I, et al. Butyrate levels in the transition from an infant- to an adult-like gut microbiota correlate with bacterial networks associated with Eubacterium Rectale and Ruminococcus Gnavus. Genes (Basel) 2020;11:1245. 16. Roswall J, Olsson LM, Kovatcheva-Datchary P, et al. Developmental trajectory of the healthy human gut microbiota during the first 5 years of life. Cell Host Microbe 2021;29:765–76e3. 17. Moeller AH, Li Y, Mpoudi Ngole E, et al. Rapid changes in the gut microbiome during human evolution. Proc Natl Acad Sci USA 2014;111:16431–5. 18. Wibowo MC, Yang Z, Borry M, et al. Reconstruction of ancient microbial genomes from the human gut. Nature 2021;594:234–9. 19. Jenmalm MC, Prescott SL. The intestinal microbiota and the child’s immune system. In: Browne PD, Claassen E, Cabana MD, eds. Microbiota in health and disease: from pregnancy to childhood. Wageningen, the Netherlands: Wageningen Academic Publishers, 2017;121–39. 20. Bager P, Wohlfahrt J, Westergaard T. Caesarean delivery and risk of atopy and allergic disease: meta-analyses. Clin Exp Allergy 2008;38:634–42. 21. Lodrup Carlsen KC, Roll S, Carlsen KH, et al. Does pet ownership in infancy lead to asthma or allergy at school age? Pooled analysis of individual participant data from 11 European birth cohorts. PLoS One 2012;7:e43214. 22. Pelucchi C, Galeone C, Bach JF, et al. Pet exposure and risk of atopic dermatitis at the pediatric age: a meta-analysis of birth cohort studies. J Allergy Clin Immunol 2013;132:616–22e7. 23. Strachan DP, Ait-Khaled N, Foliaki S, et al. Siblings, asthma, rhinoconjunctivitis and eczema: a worldwide perspective from the International Study of Asthma and Allergies in Childhood. Clin Exp Allergy 2015;45:126–36. 24. Duong QA, Pittet LF, Curtis N, et al. Antibiotic exposure and adverse long-term health outcomes in children: a systematic review and meta-analysis. J Infect 2022;85:213–300. 25. Ni J, Friedman H, Boyd BC, et al. Early antibiotic exposure and development of asthma and allergic rhinitis in childhood. BMC Pediatr 2019;19:225. 26. Campbell BE, Lodge CJ, Lowe AJ, et al. Exposure to ‘farming’ and objective markers of atopy: a systematic review and meta-analysis. Clin Exp Allergy 2015;45:744–57. 27. Genuneit J. Exposure to farming environments in childhood and asthma and wheeze in rural populations: a systematic review with meta-analysis. Pediatr Allergy Immunol 2012;23:509–18. 28. McDermott AJ, Huffnagle GB. The microbiome and regulation of mucosal immunity. Immunology 2014;142:24–31. 29. Thaiss CA, Zmora N, Levy M, et al. The microbiome and innate immunity. Nature 2016;535:65–74. 30. Shimada Y, Kinoshita M, Harada K, et al. Commensal bacteria-dependent indole production enhances epithelial barrier function in the colon. PLoS One 2013; 8:e80604. 31. Yamamoto EA, Jorgensen TN. Relationships between vitamin D, gut microbiome, and systemic autoimmunity. Front Immunol 2019;10:3141. 32. Bonakdar M, Czuba LC, Han G, et al. Gut commensals expand vitamin A metabolic capacity of the mammalian host. Cell Host Microbe 2022;30:1084–92e5. 33. Cantorna MT, Snyder L, Arora J. Vitamin A and vitamin D regulate the microbial complexity, barrier function, and the mucosal immune responses to ensure intestinal homeostasis. Crit Rev Biochem Mol Biol 2019;54:184–92. 34. Kayama H, Takeda K. Polysaccharide A of Bacteroides fragilis: actions on dendritic cells and T cells. Mol Cell 2014;54:206–7. 35. Atarashi K, Tanoue T, Shima T, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011;331:337–41. 36. Abrahamsson TR, Jakobsson HE, Andersson AF, et al. Low diversity of the gut microbiota in infants with atopic eczema. J Allergy Clin Immunol 2012;129:434–40, 440e1–2. 37. Arrieta MC, Arevalo A, Stiemsma L, et al. Associations between infant fungal and bacterial dysbiosis and childhood atopic wheeze in a nonindustrialized setting. J Allergy Clin Immunol 2018;142:424–34e10. 38. Arrieta MC, Stiemsma LT, Amenyogbe N, et al. The intestinal microbiome in early life: health and disease. Front Immunol 2014;5:427. 39. Bisgaard H, Li N, Bonnelykke K, et al. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J Allergy Clin Immunol 2011;128:646–52e1–5. 40. Chen CC, Chen KJ, Kong MS, et al. Alterations in the gut microbiotas of children with food sensitization in early life. Pediatr Allergy Immunol 2016;27:254–62. 41. Forno E, Onderdonk AB, McCracken J, et al. Diversity of the gut microbiota and eczema in early life. Clin Mol Allergy 2008;6:11. 42. Ismail IH, Oppedisano F, Joseph SJ, et al. Reduced gut microbial diversity in early life is associated with later development of eczema but not atopy in high-risk infants. Pediatr Allergy Immunol 2012;23:674–81. 43. Wang M, Karlsson C, Olsson C, et al. Reduced diversity in the early fecal microbiota of infants with atopic eczema. J Allergy Clin Immunol 2008;121:129–34. 44. Feehley T, Plunkett CH, Bao R, et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat Med 2019;25:448–53. 45. Wopereis H, Sim K, Shaw A, et al. Intestinal microbiota in infants at high risk for allergy: effects of prebiotics and role in eczema development. J Allergy Clin Immunol 2018;141:1334–42e5. 46. Arrieta MC, Stiemsma LT, Dimitriu PA, et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci Transl Med 2015;7:307ra152. 47. Stokholm J, Blaser MJ, Thorsen J, et al. Maturation of the gut microbiome and risk of asthma in childhood. Nat Commun 2018;9:141. 48. Roduit C, Frei R, Ferstl R, et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019;74:799–809. 49. Fujimura KE, Sitarik AR, Havstad S, et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat Med 2016; 22:1187–91. 50. Cahenzli J, Koller Y, Wyss M, et al. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe 2013;14:559–70. 51. Russell SL, Gold MJ, Hartmann M, et al. Early life antibiotic-driven changes in microbiota enhance susceptibility to allergic asthma. EMBO Rep 2012;13:440–7. 52. Depner M, Taft DH, Kirjavainen PV, et al. Maturation of the gut microbiome during the first year of life contributes to the protective farm effect on childhood asthma. Nat Med 2020;26:1766–75. 53. Thorburn AN, McKenzie CI, Shen S, et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat Commun 2015;6:7320. 54. West CE, Rydén P, Lundin D, et al. Gut microbiome and innate immune response patterns in IgE-associated eczema. Clin Exp Allergy 2015;45:1419–29.
Over Auteurs
Correspondentie graag richten aan: dhr. dr. J. Penders, afdeling Medische Microbiologie, Infectieziekten en Infectie Preventie, Maastricht UMC+, NUTRIM School of Nutrition and Translational Research in Metabolism, P. Debyelaan 25, 6229 HX Maastricht, tel.: 043 387 51 34, e-mailadres: j.penders@mumc.nl
Belangenconflict: geen gemeld. Financiële ondersteuning: geen gemeld.
ONTVANGEN 6 FEBRUARI 2023, GEACCEPTEERD 17 MAART 2023.
Overzicht van verschillende omgevingsfactoren in de eerste levensjaren en specifieke darmbacteriën die geassocieerd zijn met de ontwikkeling van allergische ziekten.
TABEL 1.
Overzicht van verschillende omgevingsfactoren in de eerste levensjaren en specifieke darmbacteriën die geassocieerd zijn met de ontwikkeling van allergische ziekten.
FIGUUR 1.
Overzicht van belangrijke determinanten van het microbioom in de eerste levensjaren en van een aantal manieren waarop het microbioom en microbiële metabolieten het mucosale immuunsysteem beïnvloeden.
Bron: Gecreëerd met Biorender.com
AhR=aryl-hydrocarbonreceptor, B. fragilis=Bacteroides fragilis, DC=dendritische cellen, ILC-3=type 3 aangeboren lymfoïde cellen, KKV’s=korte-ketenvetzuren, PSA=polysaccharide A, sIgA=secretoir IgA.
TABEL 2.
Overzicht van darmbacteriën die in meerdere studies geassocieerd bleken met de ontwikkeling van allergische ziekten.
Referenties
1. Al Nabhani Z, Eberl G. Imprinting of the immune system by the microbiota early in life. Mucosal Immunol 2020;13:183–9.
2. Renz H, Adkins BD, Bartfeld S, et al. The neonatal window of opportunity-early priming for life. J Allergy Clin Immunol 2018;141:1212–4.
3. Blaser MJ, Falkow S. What are the consequences of the disappearing human microbiota? Nat Rev Microbiol 2009;7:887–94.
4. Sonnenburg ED, Sonnenburg JL. The ancestral and industrialized gut microbiota and implications for human health. Nat Rev Microbiol 2019;17:383–90.
5. Rook GA, Raison CL, Lowry CA. Microbial ‘old friends’, immunoregulation and socioeconomic status. Clin Exp Immunol 2014;177:1–12.
6. Savelkoul HF. [The hygiene hypothesis: a fruitful concept in immunological research]. Ned Tijdschr Geneeskd 2006;150:2596.
7. Hornef M, Penders J. Does a prenatal bacterial microbiota exist? Mucosal Immunol 2017;10:598–601.
8. Gomez de Aguero M, Ganal-Vonarburg SC, Fuhrer T, et al. The maternal microbiota drives early postnatal innate immune development. Science 2016;351:1296–302.
9. Galazzo G, van Best N, Bervoets L, et al. Development of the microbiota and associations with birth mode, diet, and atopic disorders in a longitudinal analysis of stool samples, collected from infancy through early childhood. Gastroenterology 2020;158:1584–96.
10. Penders J, Van Best N. The development of the gut microbiota in childhood and its distortion by lifestyle changes. In: Rook GA, Lowry CA, eds. Evolution, biodiversity and a reassessment of the hygiene hypothesis. Springer Cham 2022;89:197–219.
11. Nagpal R, Tsuji H, Takahashi T, et al. Ontogenesis of the gut microbiota composition in healthy, full-term, vaginally born and breast-fed infants over the first 3 years of life: a quantitative bird’s-eye view. Front Microbiol 2017;8:1388.
12. Podlesny D, Fricke WF. Strain inheritance and neonatal gut microbiota development: a meta-analysis. Int J Med Microbiol 2021;311:151483.
13. Yassour M, Vatanen T, Siljander H, et al. Natural history of the infant gut microbiome and impact of antibiotic treatment on bacterial strain diversity and stability. Sci Transl Med 2016;8:343ra81.
14. Differding MK, Benjamin-Neelon SE, Hoyo C, et al. Timing of complementary feeding is associated with gut microbiota diversity and composition and short chain fatty acid concentrations over the first year of life. BMC Microbiol 2020;20:56.
15. Nilsen M, Madelen Saunders C, Leena Angell I, et al. Butyrate levels in the transition from an infant- to an adult-like gut microbiota correlate with bacterial networks associated with Eubacterium Rectale and Ruminococcus Gnavus. Genes (Basel) 2020;11:1245.
16. Roswall J, Olsson LM, Kovatcheva-Datchary P, et al. Developmental trajectory of the healthy human gut microbiota during the first 5 years of life. Cell Host Microbe 2021;29:765–76e3.
17. Moeller AH, Li Y, Mpoudi Ngole E, et al. Rapid changes in the gut microbiome during human evolution. Proc Natl Acad Sci USA 2014;111:16431–5.
18. Wibowo MC, Yang Z, Borry M, et al. Reconstruction of ancient microbial genomes from the human gut. Nature 2021;594:234–9.
19. Jenmalm MC, Prescott SL. The intestinal microbiota and the child’s immune system. In: Browne PD, Claassen E, Cabana MD, eds. Microbiota in health and disease: from pregnancy to childhood. Wageningen, the Netherlands: Wageningen Academic Publishers, 2017;121–39.
20. Bager P, Wohlfahrt J, Westergaard T. Caesarean delivery and risk of atopy and allergic disease: meta-analyses. Clin Exp Allergy 2008;38:634–42.
21. Lodrup Carlsen KC, Roll S, Carlsen KH, et al. Does pet ownership in infancy lead to asthma or allergy at school age? Pooled analysis of individual participant data from 11 European birth cohorts. PLoS One 2012;7:e43214.
22. Pelucchi C, Galeone C, Bach JF, et al. Pet exposure and risk of atopic dermatitis at the pediatric age: a meta-analysis of birth cohort studies. J Allergy Clin Immunol 2013;132:616–22e7.
23. Strachan DP, Ait-Khaled N, Foliaki S, et al. Siblings, asthma, rhinoconjunctivitis and eczema: a worldwide perspective from the International Study of Asthma and Allergies in Childhood. Clin Exp Allergy 2015;45:126–36.
24. Duong QA, Pittet LF, Curtis N, et al. Antibiotic exposure and adverse long-term health outcomes in children: a systematic review and meta-analysis. J Infect 2022;85:213–300.
25. Ni J, Friedman H, Boyd BC, et al. Early antibiotic exposure and development of asthma and allergic rhinitis in childhood. BMC Pediatr 2019;19:225.
26. Campbell BE, Lodge CJ, Lowe AJ, et al. Exposure to ‘farming’ and objective markers of atopy: a systematic review and meta-analysis. Clin Exp Allergy 2015;45:744–57.
27. Genuneit J. Exposure to farming environments in childhood and asthma and wheeze in rural populations: a systematic review with meta-analysis. Pediatr Allergy Immunol 2012;23:509–18.
28. McDermott AJ, Huffnagle GB. The microbiome and regulation of mucosal immunity. Immunology 2014;142:24–31.
29. Thaiss CA, Zmora N, Levy M, et al. The microbiome and innate immunity. Nature 2016;535:65–74.
30. Shimada Y, Kinoshita M, Harada K, et al. Commensal bacteria-dependent indole production enhances epithelial barrier function in the colon. PLoS One 2013; 8:e80604.
31. Yamamoto EA, Jorgensen TN. Relationships between vitamin D, gut microbiome, and systemic autoimmunity. Front Immunol 2019;10:3141.
32. Bonakdar M, Czuba LC, Han G, et al. Gut commensals expand vitamin A metabolic capacity of the mammalian host. Cell Host Microbe 2022;30:1084–92e5.
33. Cantorna MT, Snyder L, Arora J. Vitamin A and vitamin D regulate the microbial complexity, barrier function, and the mucosal immune responses to ensure intestinal homeostasis. Crit Rev Biochem Mol Biol 2019;54:184–92.
34. Kayama H, Takeda K. Polysaccharide A of Bacteroides fragilis: actions on dendritic cells and T cells. Mol Cell 2014;54:206–7.
35. Atarashi K, Tanoue T, Shima T, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011;331:337–41.
36. Abrahamsson TR, Jakobsson HE, Andersson AF, et al. Low diversity of the gut microbiota in infants with atopic eczema. J Allergy Clin Immunol 2012;129:434–40, 440e1–2.
37. Arrieta MC, Arevalo A, Stiemsma L, et al. Associations between infant fungal and bacterial dysbiosis and childhood atopic wheeze in a nonindustrialized setting. J Allergy Clin Immunol 2018;142:424–34e10.
38. Arrieta MC, Stiemsma LT, Amenyogbe N, et al. The intestinal microbiome in early life: health and disease. Front Immunol 2014;5:427.
39. Bisgaard H, Li N, Bonnelykke K, et al. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J Allergy Clin Immunol 2011;128:646–52e1–5.
40. Chen CC, Chen KJ, Kong MS, et al. Alterations in the gut microbiotas of children with food sensitization in early life. Pediatr Allergy Immunol 2016;27:254–62.
41. Forno E, Onderdonk AB, McCracken J, et al. Diversity of the gut microbiota and eczema in early life. Clin Mol Allergy 2008;6:11.
42. Ismail IH, Oppedisano F, Joseph SJ, et al. Reduced gut microbial diversity in early life is associated with later development of eczema but not atopy in high-risk infants. Pediatr Allergy Immunol 2012;23:674–81.
43. Wang M, Karlsson C, Olsson C, et al. Reduced diversity in the early fecal microbiota of infants with atopic eczema. J Allergy Clin Immunol 2008;121:129–34.
44. Feehley T, Plunkett CH, Bao R, et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat Med 2019;25:448–53.
45. Wopereis H, Sim K, Shaw A, et al. Intestinal microbiota in infants at high risk for allergy: effects of prebiotics and role in eczema development. J Allergy Clin Immunol 2018;141:1334–42e5.
46. Arrieta MC, Stiemsma LT, Dimitriu PA, et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci Transl Med 2015;7:307ra152.
47. Stokholm J, Blaser MJ, Thorsen J, et al. Maturation of the gut microbiome and risk of asthma in childhood. Nat Commun 2018;9:141.
48. Roduit C, Frei R, Ferstl R, et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019;74:799–809.
49. Fujimura KE, Sitarik AR, Havstad S, et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat Med 2016; 22:1187–91.
50. Cahenzli J, Koller Y, Wyss M, et al. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe 2013;14:559–70.
51. Russell SL, Gold MJ, Hartmann M, et al. Early life antibiotic-driven changes in microbiota enhance susceptibility to allergic asthma. EMBO Rep 2012;13:440–7.
52. Depner M, Taft DH, Kirjavainen PV, et al. Maturation of the gut microbiome during the first year of life contributes to the protective farm effect on childhood asthma. Nat Med 2020;26:1766–75.
53. Thorburn AN, McKenzie CI, Shen S, et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat Commun 2015;6:7320.
54. West CE, Rydén P, Lundin D, et al. Gut microbiome and innate immune response patterns in IgE-associated eczema. Clin Exp Allergy 2015;45:1419–29.
Correspondentie graag richten aan: dhr. dr. J. Penders, afdeling Medische Microbiologie, Infectieziekten en Infectie Preventie, Maastricht UMC+, NUTRIM School of Nutrition and Translational Research in Metabolism, P. Debyelaan 25, 6229 HX Maastricht, tel.: 043 387 51 34, e-mailadres: j.penders@mumc.nl
Belangenconflict: geen gemeld. Financiële ondersteuning: geen gemeld.
Om de beste ervaringen te bieden, gebruiken wij en onze partners technologieën zoals cookies om informatie over het apparaat op te slaan en/of te openen. Toestemming voor deze technologieën stelt ons en onze partners in staat om persoonlijke gegevens zoals surfgedrag of unieke ID's op deze site te verwerken en om gepersonaliseerde en niet-gepersonalisserde advertenties te tonen. Indien u geen toestemming geeft of deze intrekt, kan dit invloed hebben op bepaalde functies.
Klik hieronder om in te stemmen met het bovenstaande of om specifieke keuzes te maken. Uw keuzes zullen alleen worden toegepast op deze site. U kunt uw instellingen te allen tijde wijzigen, inclusief het intrekken van uw toestemming, door gebruik te maken van de knoppen op het Cookiebeleid of door te klikken op de knop 'Toestemming beheren' onderaan het scherm.
Functioneel
Altijd actief
De technische opslag of toegang is strikt noodzakelijk voor het legitieme doel het gebruik mogelijk te maken van een specifieke dienst waarom de abonnee of gebruiker uitdrukkelijk heeft gevraagd, of met als enig doel de uitvoering van de transmissie van een communicatie over een elektronisch communicatienetwerk.
Voorkeuren
De technische opslag of toegang is noodzakelijk voor het legitieme doel voorkeuren op te slaan die niet door de abonnee of gebruiker zijn aangevraagd.
Statistieken
De technische opslag of toegang die uitsluitend voor statistische doeleinden wordt gebruikt.De technische opslag of toegang die uitsluitend wordt gebruikt voor anonieme statistische doeleinden. Zonder dagvaarding, vrijwillige naleving door uw Internet Service Provider, of aanvullende gegevens van een derde partij, kan informatie die alleen voor dit doel wordt opgeslagen of opgehaald gewoonlijk niet worden gebruikt om u te identificeren.
Marketing
De technische opslag of toegang is nodig om gebruikersprofielen op te stellen voor het verzenden van reclame, of om de gebruiker op een site of over verschillende sites te volgen voor soortgelijke marketingdoeleinden.